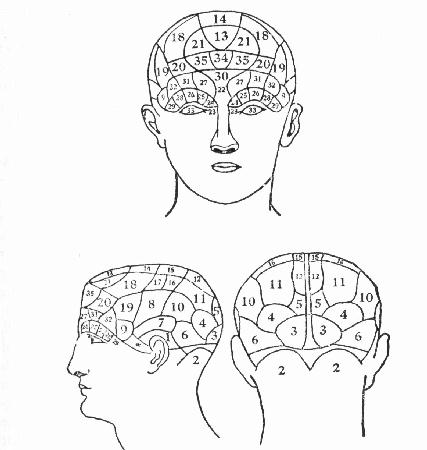
 Figure 1. From Spurzheim (1834).
Figure 1. From Spurzheim (1834).Explanations of activities of living systems in the life sciences have often taken the form of mechanistic explanations in which the activity of the system is identified with a component of the system. The overall activity is decomposed into sub-functions, which are then localized within the responsible component (Bechtel and Richardson, 1993). Mental activities have been no exception. The brain (including the central nervous system) has long been taken to be the responsible organ of the body for mental activities, and the task has been to decompose mental functions and identify the resulting component functions with parts of the brain. In many respects, this effort was the product of adopting the vision of Franz Joseph Gall.
The pursuit of mechanistic explanations via decomposition and localization has always been controversial, both for biological functions generally and mental activities in particular. In biology the opposition has generally been termed vitalism. While vitalists such as Bichat and Pasteur were often right in rejecting the overly simplistic mechanistic accounts of their contemporaries, they themselves generally failed to advance the effort to explain functions since they offered little in the way of a positive methodology. Much the same has been true with mental activities. Some of the opponents of mechanism (e.g., Flourens) have been overtly dualists and have denied that knowledge of the brain could contribute to the development of explanations of mental activities. Others (e.g., Lashley) have acknowledged that the brain was responsible for mental activities, but simply resisted any attempt to differentiate cognitive functions across brain structures. Some contemporary advocates of dynamical systems theory seem to fall in the latter camp (van Gelder, van Orden), although, unlike early opponents of mechanistic explanation, these theorists do offer an alternative strategy. We shall, at the end, argue that their opposition to decomposition and localization is misguided.
Despite the challenge of dynamical systems theory, modern cognitive neuroscience, the most sophisticated attempt yet to link mental functions with neural structures, has embraced the quest for mechanistic explanation through decomposition and localization. Thus, part of the legacy of Gall is very much still alive. However, there are different strands in this legacy. Part of our goal is to differentiate three of these strands. One, which we link with Fodor's proposal for modularity, follows Gall himself in seeking direct neural instantiation of mental functions. A second strand, more prevalent in philosophical accounts of mind-brain relations than actual empirical inquiries, but implicit in psychological research which is cut off from neuroscience, recognizes the need to localize functions at a finer grain, but adopts a top-down perspective in which decomposition precedes localization. The third construes decomposition and localization as each informing the other. One element of this view is the claim that brain regions are not independently demarcated, but are in part identified through the process of assigning functions to them. The second element, and the one to which we will devote primary attention, is that information gleaned from neural studies of component structures often leads to fundamental revisions in functional decompositions of cognitive performance. It is this (neo) Gallean strand that we see as most common in cognitive neuroscience, and which we will be principally concerned to explicate through examples.
1. Gall and the Program of Direct Localization
Gall is widely known for his proposal that mental faculties are localized on the surface of the brain. Gall was not, however, the first to propose that regions of the brain had distinctive mental functions. Several investigators in the 18th century had proposed localization of either sensory motor functions or of psychological capacities. However, the legacy of Albrecht von Haller, who contended that all neural tissue was highly irritable, and thus provided a sensorium commune, and who rejected any attempt at localization of specific functions in the brain, dominated the research landscape in the 18th and early 19th century (see Clarke and Jacyna, 1987).
While Gall's organology (more widely known as phrenology, the term supplied by his collaborator Spurzheim in 1818) is largely ridiculed, in part because of the quackery the surrounded its practices, it was based on a number of assumptions, some plausible but mistaken, some more radical but ultimately more influential. One of the mistaken assumptions was that the skull fit closely to the brain so that the shape of the skull provided a reliable indicator of the shape of the brain. Another assumption, which has earlier roots and enduring legacy, was that there were distinct mental capacities or faculties and that individuals differ in these faculties. These faculties characterize the different abilities or propensities to action, upon which individuals might differ, such as verbal memory (the first ability Gall focused on), destructiveness, and physical love. Gall identified 27 such faculties: Spurzheim and others later expanded his list. Assumptions like this persist today in such forms as Chomsky's claim that there is a language faculty (Chomsky, 1980, Pinker, 1994). Yet a third assumption, and a radical one given von Haller's influence, was that these faculties were localized in distinct parts of the brain. Fourth was the assumption that greater development in a localized brain area corresponded with larger size. This assumption, as plausible as it might have seemed, has been discredited. But it has been replaced with a conceptually related assumption that greater activity in a brain area corresponds with greater metabolism (e.g., blood flow).
Figure 1. From Spurzheim (1834).
Given these assumptions, the motivation for seeking correlations between skull shapes and distinct psychological faculties, and using this to infer the function of underlying brain structures seems well motivated. The actual linking of particular faculties with skull shapes was an empirical enterprise, and here Gall's work is most suspect. Gall tended to focus on examples of individuals with special development or deficit in particular faculties and protrusions or indentations in parts of their cranium that corresponded, in his scheme, to these faculties. From these cases he advanced his famous maps of functions (Figure 1). But what is more important for us is the general view he defended, in which specific brain regions were responsible for specific psychological capacities:
When we see that nature follows such a course, how can it still be doubted that each part of the brain has different functions to fulfill, and that as a consequence, the brain of man and animals must be composed of as many special organs as the man or animal has distinct moral or intellectual faculties, inclinations, and aptitudes for work (Gall and Spurzheim, 1812, p. 254).
Gall's legacy was invoked by many neuroscientists later in the 19th century as they began to identify regions of the brain with specific mental activities. These theorists sharply distinguished Gall's assumptions about how the cranium mapped underlying brain structure from his contention that different brain areas carried out different mental processes. They then developed new tools, such as linking deficits to lesions or direct electrical stimulation of the brain, to localize functions in the brain. But by the 20th century, Gall came to be held in far lower repute until he was "rehabilitated" by Fodor (1983). Fodor contended that a good part of the mind had the character of Gall's faculties; he identified these areas as modules and claimed that they were responsible for processing particular contents. But Fodor went far beyond Gall in emphasizing the encapsulation of these modules--each module had to rely on its own source of information and its operation could not be penetrated by other information the subject possessed. Fodor's account departs from Gall's in that he only identified such modules in specific sensory input domains and in language processing and he viewed these modules not as directly controlling particular behaviors, but as supplying input to more central cognitive processes which are responsible for higher level reasoning and operate in a more holistic manner. One kind of information that cannot penetrate the input modules, then, is information held by central cognition. Fodor also identified a number of other characteristics of information processing within modules: such processing is very fast, automatic, and mandatory (when the input is received, it is processed). He also contended that such modules are likely to be hardwired into the cognitive system. Accordingly, they would presumably be systems that are physically identifiable in the brain, although Fodor does not emphasize the physical localization.
One important point to note about both Gall and Fodor is that for both of them the analysis of function comes prior to the analysis of structure. Gall (using "physiology" to refer to function) wrote:
The physiology of the brain need not be contradictory to the anatomy of the brain; its anatomy must become the support of its physiology. But physiology has been found independently of anatomy. Nothing whatever in brain physiology has conflicted with an anatomical fact; nothing has been interpreted by structure or by the arrangement of parts of the brain; brain anatomy has served only to confirm physiological discoveries. (1825, p. 118)
For Fodor, likewise, functions are identified autonomously from their brain realization. The primary criteria for identifying modules are their functions--each module is specific to a given sense or to language.
2. Top-Down Complex Localization
In linking his modularity thesis to Gall's organology, Fodor characterizes the faculties or modules as "vertical" and distinguishes them from another sort that are common in cognitive theories, which he calls "horizontal." The difference lies in the fact that Gall's and Fodor's faculties are responsible for a given content domain whereas horizontal faculties operate across content domains. Short-term memory, if there turns out to be such a system, stores information on a variety of contents. Moreover, in carrying out any given task, it is assumed to interact with other cognitive capacities. In Fodor's perspective, these horizontal faculties belong to central cognition, and he is deeply pessimistic about the ability of cognitive science to make any progress in explaining these capacities. Cognitive science nonetheless has generally taken these horizontal faculties to be its principal focus and has tried to develop a number of tools for demonstrating the existence of particular processing components and determining how they interact with each other. (And, as we shall see, cognitive neuroscience offers consolation to the worry that central cognition cannot be mechanistically explained.)
Identifying horizontal modules requires going beyond simple localization; it requires decomposing a given task into subtasks. The need for decomposition in order to explain is independent of whether an initial direct localization is correct. If it is, then the decomposition occurs within the module. But a common fate of proposed direct localizations in the life sciences, including cognitive neuroscience, is that the structure in which a function is first localized turns out to be only one contributor to the performance of the task. For example, shortly after Broca (1861) identified a region in the brain as the source of language deficits and thus as potentially responsible for articulate language, Wernicke (1874) identified a second brain region that also produced language deficits, although of a different sort, and proposed a model in which different areas of the brain contributed in different ways to language functioning. Such a differentiation of tasks is a step toward developing a functional decomposition of an overall function into sub-tasks or component functions, which then together accomplish the activity. It is these functions which a more adequate explanatory account will hope to localize in the brain. A central methodological question concerns how these component functions are to be identified. In Wernicke's case (and in the case of Geschwind, 1974, who carried on Wernicke's tradition), brain lesions have been the primary guide to functional decomposition. But brain lesions have played only a minor role in the decompositions developed in cognitive science.
One approach is to think that functional decompositions can be developed a priori by asking what activities must be performed in order to perform the overall task. Such an approach is common in engineering: one designs a machine to carry out a task by first identifying component tasks, then identifying component tasks within these tasks, and so on, until one reaches a point where the tasks invoked in the analysis correspond to the capabilities of known mechanical devices. Then one puts the devices together in such manner as to interact appropriately to perform the task. This approach is exemplified in designing computer programs: the programmer must analyze the overall performance that is sought into component activities until these activities correspond to the primitive operations of one's programming language or to already developed programming modules. Such an approach has been followed by many investigators in artificial intelligence. A common result is that initial proposals as to how a given task should be decomposed turn out to be inadequate to account for the way the task is usually performed, and researchers need to develop alternative models, including new modeling tools (e.g., scripts and frames) and new modeling frameworks (e.g., production systems and case based reasoning systems). In all these cases, though, the guide to developing new tools and frameworks has come from considerations of the task that needs to be performed (e.g., making a medical diagnosis or reading a newspaper story) and means-end reasoning about that task.
Another approach to identifying functional components is through behavioral experimentation. The goal here is to find evidence in the behavior of the mind/brain about how it breaks down tasks. The challenge in such research is to find a variable to measure that might point to the differentiation of tasks. Two of the most widely invoked are reaction times and error measures. In reaction time studies one measures the amount of time it takes one to perform a given task. The problem is that one must measure a whole activity, which involves sensory processing, whatever central activities are involved, and motor processing. To get at the amount of time involved in a specific cognitive task, one generally has to do comparative studies. One of the first approaches to using reaction times to identify cognitive processes was Donders' (1868/1969) use of the subtraction method in which one devised two tasks in which one could be envisaged as adding an additional step to what was required in the other task. The time difference in the performance of the two tasks was then a measure of how long the additional task required. There are at least two problems with this approach. First, it requires a proposal for a task decomposition before one can set up the substraction. Second, it assumes that performing the additional task simply adds to the other operations and does not occur simultaneously with them. Saul Sternberg (1967) developed a procedure, known as the additive factors method, to overcome this later problem. In this procedure one devises procedures to alter different variables that are presumed to affect different component tasks. If the changes in reaction times resulting from manipulating two variables are additive, then the affected component tasks are revealed to be independent, even if they occur simultaneously. If, on the other hand, the change in reaction time from manipulating two variables is not equal to the sum of changes for manipulating the variables independently (this is called an interaction), then the component tasks are not independent. A related way of demonstrating whether two components are independent is the interference paradigm. If requiring a subject to perform a second task while performing an initial task leads to increased reaction times, this provides evidence that a common component is employed in both tasks.
While all of these methods begin with an hypothesis about the decomposition of the task, it should be emphasized that the additive factors method and the interference paradigm provide a means for possibly falsifying the initial hypothesis and so guiding the quest for a more adequate decomposition. What is important for our purposes, though, is to note that they start on the behavioral side, using behavioral data to test and revise task decompositions. The task decomposition is set, therefore, before one might begin to consider neural evidence.
So far we have focused on top-down decomposition in artificial intelligence and cognitive psychology. But the program has also been endorsed in philosophy of mind. The dominant philosophical account of mind of the past 30 years is functionalism, which holds that mental activities are to be understood in terms of their interaction with other mental activities, not their physical realization. One strand of functionalism, homuncular functionalism, advocates a multi-stage decomposition of cognitive performance, usually first framed in intentional terms, but eventually reaching a point at which a task is sufficiently simple that one can envisage a mechanistic system performing it. Dennett characterizes this as the approach of AI researchers:
The AI researcher starts with an intentionally characterized problem (e.g., how can I get a computer to understand questions of English?), breaks it down into sub-problems that are also intentionally characterized (e.g., how do I get the computer to recognize questions, distinguish subjects from predicates, ignore irrelevant parsings?) and then breaks these problems down still further until he reaches problem or task descriptions that are obviously mechanistic (1978, p. 80).
One stream of functionalism takes the emphasis on function to show that reduction of psychological states to brain states is not possible, largely because psychological states can be realized in very different types of brain states (Putnam, 1975, Fodor, 1987). Accordingly, that strand of functionalism often denies that knowledge of the brain can be informative of psychological processes. Homuncular functionalism, as Lycan (1981) makes clear, does not endorse such a chasm between mind and brain, and indeed is compatible with the classical theory of the identity of mind and brain. Closely identified with the identity theory is a model of reduction, according to which psychological processes are identified with neural processes and psychological laws are derived from laws of neuroscience. The resulting picture is nicely described by Chalmers:
The paradigm of reductive explanation via functional analysis works beautifully in most areas of cognitive science, at least in principle . . . most nonphenomenal mental concepts can be analyzed functionally. Psychological states are characterizable in terms of the causal role they play. To explain these states, we explain how the relevant causation is performed . . . In principle, one can do this by giving an account of the underlying neurophysiology. If we explain how certain neurophysiological states are responsible for the performance of the functions in question, then we have explained the psychological state. We need not always descend to the neurophysiological level, however. We can frequently explain some aspect of mentality by exhibiting an appropriate cognitive model - that is, by exhibiting the details of the abstract causal organization of a system whose mechanisms are sufficient to perform the relevant functions, without specifying the physicochemical substrate in which this causal organization is implemented. In this way, we give a how-possibly explanation of a given aspect of psychology, in that we have shown how the appropriate causal mechanisms might support the relevant mental processes. If we are interested in explaining the mental states of an actual organism or type of organism (e.g., learning in humans, as opposed to the possibility of learning in general), this sort of explanation must be supplemented with a demonstration that the causal organization of the model mirrors the causal organization of the organism in question (Chalmers 1996, p.46).
The approach of AI, cognitive psychology, and homuncular functionalism in philosophy that we have characterized in this section emphasizes a top-down direction to the development of mechanistic theories of mind: the decomposition is first developed by analyzing behavior, and perhaps subsequently connected to underlying neural processes. There are some vocal opponents of the picture of functional decomposition first, structural localization second, such as Paul and Patricia Churchland. In their extreme eliminativist mode, they recommend ignoring the functional decomposition of cognitive capacities in favor of an approach that focuses solely on the brain. (As McCauley, 1996, argues, based on a number of texts, this is not the only position that the Churchlands adopt. In other contexts, they embrace the sort of interactive approach that we discuss in the next section.) Insofar as they endorse eliminating not just folk psychology, but also the functional analyses of cognitive psychology and cognitive science, when reduction fails, they seem to endorse the view that if such functional analyses are to play any role in a proper science, it must be that of providing a prior decomposition that is to be completed before neuroscience takes over to localize functions. The eliminativist claim is that neuroscience won't find any components carrying out these functions, and that the whole functional analysis will be eliminated.
3. Interactive Decomposition and Localization
Cognitive neuroscience, both in its late 19th and early 20th century development, and in its current pursuit, supports a different perspective on the functional decomposition and neural localization of mental function. The identification of neural structures, the very 'carving' of the brain, does not proceed independently of an assumed functional decomposition of cognitive capacity and the functional decomposition of cognitive capacity is informed by neuroanatomical evidence (Mundale and Bechtel, 1996). These features of the methodology of contemporary cognitive neuroscience, exemplified in the cases we discuss, set it apart from the Gallean direct localizationist tradition and the top-down approach to reductionism found in much contemporary philosophy of mind. The complex interplay between cognitive, functional decomposition and neural localization that we highlight suggests that the study of complex systems like the brain is an inherently inter-disciplinary endeavor, involving the close cooperation of many different theoretical perspectives, rather than their independence in a strict division of labor. To illustrate the nature of this dynamic interaction, we will analyze two lines of research in contemporary cognitive neuroscience. The first began with work on visual processing in monkey brains that pointed to a rather surprising decomposition between two visual processing systems, one primarily concerned with identifying what an object is and the other with determining where it is. As we shall see, once this decomposition was suggested from research on the brain, it has led to new threads of research on the cognitive side, both in terms of computer modeling of cognition and behavioral research. The second line of research examines developments in the study of attention. Here again information stemming from the brain has prompted revisions in the decomposition of cognitive function.
a. Decomposition and Localization in the Visual Processing System
The phenomenology of seeing is one of direct access to nearly all aspects of the visual field. We are not aware of this task being decomposed into subtasks. But when one asks how to get a machine, biological or silicon based, to do it, it is immediately obvious that this is not a simple, direct process. We are not aware of any physical process that directly transforms sensory input into a perception of the world in which one recognizes objects and their significance, where they are, and how they are moving. Since perceiving is not a simple physical process, if we are to understand it or replicate it, we need to decompose it into subtasks that are more directly physically realizable. This task has been taken up in a practical way in computer vision. Here one starts with a characterization of the nature of the information received at the sensory threshold [1] and then tries to devise algorithms which operate on this information until a perception of a three dimensional world is built up. One of the more celebrated efforts of this sort is that of Marr (1982). This type of approach decomposes the process in a nearly totally top-down fashion: one employs means-ends analysis in the attempt to figure out how information might be manipulated to get from the assumed input to the assumed target output.
Neuroscience offers a different opportunity for decomposition, and one that has already proven richly rewarding in the approximately quarter century that it has been applied. The difference lies in the tools for developing the task decomposition. Instead of relying on one's abilities in means-end analysis to come up with a decomposition, neuroscience offers a variety of empirical tools: neural-anatomical studies of cell types and patterns of connectivity, recordings from individual neurons, studies of deficits when specific neural structures are destroyed, and most recently imaging of activity patterns in actively functioning brains. None of these alone offers a simple and foolproof route to determining how the brain decomposes the task of visual perception, and where the processes are localized, but together they offer a set of tools for constraining the decomposition and localization.
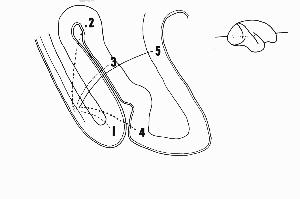 Figure 2. From Zeki (1976).
Figure 2. From Zeki (1976).
It has long been recognized that the key cortical areas for initiating visual processing lie in the rear of the brain in the occipital lobe, more specifically, in Brodmann's area 17 or V1. Neuroanatomical studies in the 1960s showed five different pathways from V1 into Brodmann's area 18, or prestriate cortex: to V2 (2), V3 (3) , superior temporal sulcus (5), V4 (3), and V4a (4) (Craig, 1969). (Figure 2) This suggested the possibility of different types of processing along each of these pathways. By providing different visual stimuli and recording from cells in each of these areas, researchers such as Zeki (1976) discovered that cells in different regions of prestriate cortex responded to different types of information--e.g., those in V2 to binocular disparity, those in V4 to specific colors, while those on the superior temporal sulcus responded to motion.
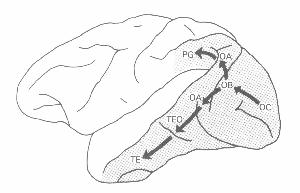 Figure 3. From Mishkin et al. (1983).
Figure 3. From Mishkin et al. (1983).
One of the most heralded findings of modern neuroscience, and we would suggest most surprising from the point of view of attempts at top-down decomposition of cognitive processing, arose from tracing the processing beyond the prestriate cortex areas. Mishkin, Ungerleider, and Marko (1983), relying largely on lesion studies in monkeys (the first study in this series was Pohl, 1973), differentiated two main routes for processing visual information. (Figure 3) From prestriate cortex, one route proceeds ventrally into posterior temporal cortex along the inferior longitudinal fascilus to areas TEO and TE. Based on the fact that lesions in the posterior temporal area result in loss of pattern discrimination and that lesions in TE in particular result in failure to recognize previously presented objects, Mishkin et al. assigned to this pathway the task of analyzing "the physical properties of a visual object (such as its size, color, texture and shape)" (p. 20). Since there are neural pathways between TE and limbic structures in the temporal lobe and ventral portions of the frontal cortex, Mishkin et al. further claim that this area figures in assigning meaning to visual stimuli. One feature of neurons in the ventral pathway is that as one proceeds further, the cells have increasingly large receptive fields, suggesting that they become responsible for recognizing objects independently of where they appear in the visual field. Performing a location independent identification, they contend, requires discarding any location information:
It is significant that by virtue of the extremely large visual receptive fields of inferior temporal neurons this area seems to provide the neural basis for the phenomenon of stimulus equivalence across retinal translation; i.e. the ability to recognize a stimulus as the same, regardless of its position in the visual field. But a necessary consequence of this mechanism for stimulus equivalence is that within the occipitotemporal pathway itself there is a loss of information about the visual location of the object being identified (p. 21).
The other route proceeds from prestriate cortex dorsally into the posterior parietal cortex. Lesions in the posterior parietal cortex in monkeys result in failure to be able to select a response location on the basis of a visual landmark, suggesting that this pathway figures in "perception of spatial relations among objects, and not in their intrinsic qualities" (p. 21). Further studies revealed that parietal area PG is responsive to tactile input as well as visual, suggesting that it processes polysensory spatial information. Unlike the ventral pathway, this parietal pathway is limited to input from the contralateral part of the visual field. Moreover, whereas the temporal pathway is primarily sensitive to foveal areas, the parietal pathway is equally sensitive to foveal and peripheral areas.
The two systems differentiated by Mishkin et al. quickly became known popularly as the "What" and "Where" systems. Further research revealed that the two pathways diverge even before V1 and employ different cells in V1. The retina projects to V1 via the Lateral Geniculate Nucleus (LGN) and even in the LGN there are two steams, a parvocellular (P) and a magnocellular (M) stream. [2] These two streams receive projections from different retinal ganglia and project into different cells in V1, with the P pathway then projecting into inferotemporal cortex (the What system) and the M pathway projecting into parietal cortex (Where system) (Maunsell and Newsome, 1987). The original work differentiating the two systems used lesion and deficit studies, but it was complemented by blood flow tracer studies with radioactively labeled deoxyglucose (Macko, Kennedy, Sokoloff, and Mishkin, 1981). These revealed one pathway from V1 to V2 (OB in Figure 3), to V3 and V4 (both in ventral OA in Figure 3), and from V4 to both TEO and TE, and another that begins with projections from both V1 and V2 to MT (mostly within dorsal OA), and from there to four areas in the upper superior temporal and intraparietal sulci (which is within PG).
The advent of PET provided a way of demonstrating functionally distinct pathways in normal humans, and Haxby, Grady, Horwitz, Ungerleider, Mishkin, Carson, Herscovitch, Schapiro, and Rapoport (1991) proceeded to take advantage of this new technology. In PET studies, blood flow is measured (indirectly) in different tasks; the resulting patterns produced for each task can then be subtracted from one another so as to reveal areas of the brain that are active in one but not the other task. Haxby et al. used a face matching and a dot matching task as the target tasks, and subtracted from the resulting images the images produced in a sensorimotor control task (this is the same substraction methodology mentioned above in the context of reaction time studies). Their major finding, in accord with the What/Where distinction, was that
The face-matching task caused rCBF [regional cerebral blood flow] increases in a broad region extending from the occipital pole to posterior temporal cortex in all subjects. The dot-matching tasks caused rCBF increases in superior parietal cortex in 8 of 11 subjects and rCBF increases in superior parietal cortex in 10 of 11 subjects. The lateral occipital area activated during spatial visual processing overlapped with the areas activated during face matching but tended to be closer to the occipitoparietal border and did not include the more anterior and superior occipitotemporal region (1623).
While the What/Where distinction arose first in neuroscience, it has subsequently influenced research in computational and behavioral areas of cognitive science. In computational modeling, it helped foster inquiry into modular networks by connectionists. Early connectionist models tended to be layered networks in which each unit in one layer was connected to every unit in preceding and succeeding layers. In modular networks, different tasks are performed by different subnetworks, thus introducing a task decomposition. One sort of modular design for neural networks involves having several discrete networks receive input from the same source, but with different networks performing different tasks with this input. Each network will respond to every input, but either the networks can be trained to perform different tasks on the input by having separate response units, or a gating network can regulate which network will have access to the output units for a given task, and consequently learn from its performance on that task. In a first demonstration of the potential of modular networks to solve cognitive tasks, Rueckl, Cove, and Kosslyn trained a network with separate sets of hidden and output units to identify the object in one subnetwork and specify where it was in another. The input was a 5x5 "retina" in which any one of nine different object patterns could occupy one of the nine different 3x3 matrices within the retina (in Figure 4, for example, a "T" is in the central matrix). They showed that the network learned much more quickly if one used separate sets of the hidden units and allowed one set to feed each set of output units than if there were a common set of hidden units.
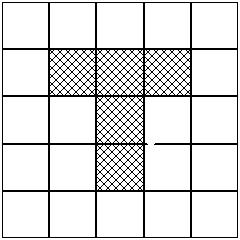 Figure 4
Figure 4
Jacobs, Jordan, and Barto (1991) extended the demonstration to show that a network could learn to subdivide these tasks between subnetworks. They employed a gating network, which learned to control which subnetwork had access to the output units and received the learning signal (Figure 5).
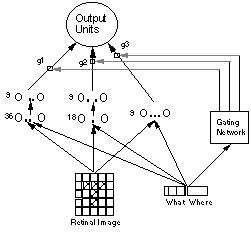 Figure 5.
Figure 5.
On a given task, the network was to specify either the location or the name of the object. The subnetworks each had a different configuration (one had no hidden layer, one had 18 hidden units, and the third had 36 hidden units). Through backpropagation learning in both the modular subnetworks and the gating network, the system learned in 50 epochs to divide the tasks so that the network with no hidden units performed the where task and the network with 36 hidden units performed the what task. This is, in fact, an optimal distribution since the where task is linearly separable and thus efficiently handled by a network without hidden units, whereas the what task requires hidden units. As a comparison, Jacobs et al. attempted to train a single network to handle both tasks, but such a network never learned as well, and was subject to catastrophic interference in which learning one task interfered with the ability to perform the other task. What this suggests is that the decomposition employed by the brain may be computationally efficient.
The What/Where distinction has also begun to have an impact in psychology. One of the first invocations of it was by Neisser (1989), who invoked it to suggest how Gibsonian ecological psychology and computational cognitive psychology might be brought together. He contended that Gibson was most concerned with perceiving aspects of the environmental layout and that this might be determined directly. Jacobs et al.'s demonstration that where decisions require only a two-layer network and hence no intermediate steps might provide some sense to this directness claim, although probably not the one Gibson and Neisser were concerned with. The what system, on the other hand, may have to carry out more complex operations, making a decomposition of this task into multiple inferential steps more plausible. Until recently, however, there was little purely psychological evidence for the What/Where distinction. Indeed, evidence in the speed of processing literature suggested that there was only one factor that accounted for a subject's speed of processing on a wide variety of cognitive tasks. Hale and Jansen (1994), for example, showed that on a variety of cognitive tasks (choice reaction time, letter classification, visual search, abstract matching, line-length discrimination, mental rotation, and mental folding), an individual subject's reaction times were a linear function of the mean reaction times of an entire group. Thus, a given individual might have reaction times slower or faster than the whole group, but this speed up or slow down would carry across all tasks studied. This provided evidence of a single "global processing-time coefficient [that] characterizes the speed of all of an individual's basic cognitive processes" (388). This is not what one would expect if there were different cognitive systems performing these tasks. However, the tasks in Hale and Jansen's study were not chosen so as to differentiate spatial location tasks from object identification tasks. More recently, Hale and her colleagues (Hale, Chen, Myerson, & Simon, 1996) have devised tasks to differentially emphasize spatial location determination and object identification procedures. In this new study, they have found subjects with differential advantage either on what tasks or where tasks.
Our major contention is that in this case, information drawn from the brain has played a major role in developing the task decomposition. The what/where distinction did not arise from top-down theoretical considerations, but from differentiating neural components that did different tasks. Once identified, though, the decomposition can be employed in all the relevant disciplines. Thus, we have seen how it has fed back into computational modeling and into psychological experimentation.
Unfortunately, we must note that while the decomposition and localization of visual processing areas, and the spin offs into computational modeling and experimental psychology, that we have discussed so far present an intellectually coherent and satisfying story, nature may turn out to be more complicated. The details of the neuroanatomy of the visual cortex of the macaque indicate far more communication between processing channels than the two-pathway model acknowledges. Van Essen and his colleagues (Felleman and van Essen, 1991; van Essen and DeYoe, 1995), using a
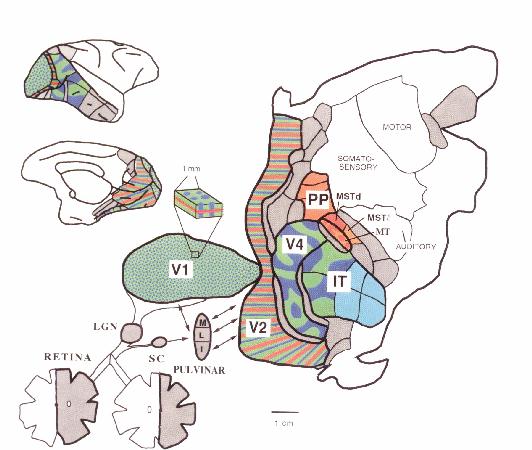 Fig. 6: From van Essen and Gallant (1994), p. 2.
Fig. 6: From van Essen and Gallant (1994), p. 2.
variety of techniques to flatten and map the macaque visual cortex,
identify 32 distinct areas (Figure 6). [3] They have
also shown that these areas on average receive 10 inputs and send 10
outputs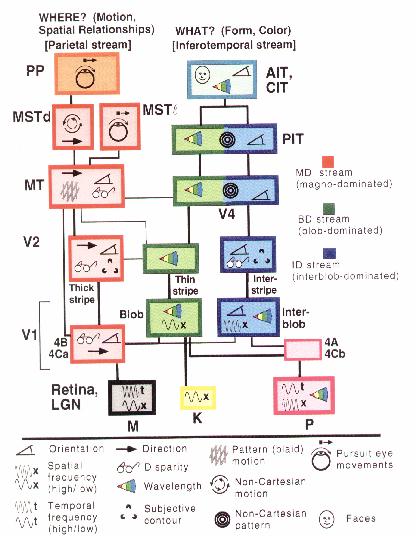 Figure 7. From van Essen and Gallant (1994), p. 3.
Figure 7. From van Essen and Gallant (1994), p. 3.
to other areas so that overall nearly one third of the possible corticocortical connections are realized. (Some of these pathways may be relatively weak, as they are "only faintly detectable using the most sensitive available technique", van Essen and DeYoe, 1995, p. 386). Moreover, these connections tend to be reciprocal. [4] Cross talk between the different pathways begins already in V1. The M stream cells in V1 receive inputs from both the M and P steam, although inputs through the M stream predominate. Moreover, axons project across the 4C/4C boundary (see Figure 7), and there are further such projections on up the processing pathways. From anatomical studies alone one cannot determine the functional significance of different projections; more direct functional evidence can be gathered by selectively inactivating a particular input stream and observing the effects on cells downstream. The fact that even after inactivating P-layer cells in LGN, cells in the blob area of V1 can still be driven indicates that they are receiving functional inputs from the M and K streams (Nealey, Ferrera, and Maunsell, 1991). Both the blob and the interblob areas (the latter are primarily driven by P cells in LGN) of V1 project to distinct parts of V4 and the posterior inferior temporal cortex, although V4 also receives input from the cells in V3, part of the M stream.
If the contentions of van Essen and his colleagues are correct, then the mechanistic account of visual processing will turn out to be more complex than expected. This, however, is the norm in science. The first accounts of biochemical processes such a fermentation and oxidation proposed moderately simple, linear chains of reactions. Subsequent research resulted in the discovery of significant nonlinearities. These, however, did not require repudiating the attempt to decompose biological processes, but the development of more complex accounts of what the contributing processes are and how they are integrated (Bechtel, 1986, Bechtel and Richardson, 1993). We suspect that much the same is now happening in understanding of visual processing. As research continues, decomposition and localization will continue to interact with each other. In some cases decompositions developed from focusing on the task and carrying out means-end reasoning will provide the basis for identifying physical components and their functions. In other cases, the discovery of the components may lead in the process of developing the decomposition.
b. Decomposition and Localization of Attentional Mechanisms
Attention has been a topic of consideration in cognitive theorizing since the cognitive revolution, but research here too has recently benefited from brain based investigation. Our discussion will focus on three models of attention. The first was introduced by Broadbent (1958) and others primarily on the basis of behavioral evidence that suggested the existence of a limited cognitive resource, or 'bottle-neck,' that stimulus information competes for, somewhere between perception and motoric response. The second model was introduced by Michael Posner on the basis of behavioral and neural evidence. According to the second model there are three attentional systems, a visuo-spatial, posterior network, a non-modality-specific, top-down, anterior network, and a vigilance network. The posterior system is conceptualized as a 'spotlight' that illuminates different locations in the visual field. The third model is Desimone and Duncan's attentional template, biased competition model, in which different processing streams compete for the control of overt behavioral response, and receive biases from an attentional template according to their relevance to the demands of a task.
Attentional effects are measured behaviorally by comparing reaction
times and error scores on tasks under different conditions. Differences
in performance are taken to indicate that under certain conditions human
information processing resources are more taxed than under other conditions.
Thus, in one type of task, the visual search task, two conditions are compared.
In both conditions, there is an array of objects or shapes that must be
searched in order to discover some target that is  Figure 7.
Figure 7.
pre-specified in terms of a unique feature or set of features. In the
first condition, the target is pre-specified in terms of an obviously distinguishing
feature. For example, the array may consist of a set of green shapes and
one red shape, and the subject may be asked to identify the red shape (Figure
7). Reaction time in this condition does not increase with the number of
green shapes, or distractors, in the array. In the second condition, the
target is pre-specified in terms of a non-obvious feature or set of features,
like a conjunction of features. For example, the array may consist of different
shapes of various colors, and the subject may be asked to identify the
yellow triangle . Figure 8.
Figure 8.
(Figure 8). Relative to this array, this conjunction of properties is non- obvious. The subject must engage in controlled, or attention-demanding processing, to discover the target. It is for this reason that reaction time in this condition increases with the number of distractors, i.e., colored shapes, in the array.
The limited capacity model explains the different performance in these two conditions in the following way. In the first condition, a relatively unlimited, pre-attentive perceptual capacity scans the array in parallel, and selects the target automatically for further processing by the limited attentional resource, and eventually for motoric response. Because the array is scanned in parallel by a relatively unlimited, pre-attentive cognitive resource, the number of distractors has little or no effect on reaction time. In the second condition, the limited attentional resource must be used in a controlled, serial search for the target. Each item in the array must be selected for processing by the limited attentional resource, in sequence, until the target is found. It is for this reason that reaction time increases with the number of items to be processed.
The understanding of such results in terms of selection by a limited attentional resource suggests a very definite research program. Attention is functionally analyzed as a processing 'bottle-neck' or limited capacity resource that selects from competing stimulus information represented automatically and in parallel with pre-attentive perceptual resources. Such a model raises the following questions. When in the processing of perceptual input, and on what basis, is information selected for processing by the limited attentional resource? What type of processing, e.g., semantic, formal, etc., is the limited attentional resource required for? What are the relationships and differences between controlled and automatic processing? These functional questions about the types of processing that distinguish the limited attentional resource from less limited pre-attentive resources suggest corresponding questions about the localization of the relevant cognitive functions. Where in the brain are limited attentional resources found? Where in the brain does pre-attentive, automatic, parallel processing occur? How does selection among information competing for attention occur in the brain?
Michael Posner is one of the first cognitive psychologists to bring
brain research to bear on the study of attention and, as we shall see,
in his hands brain studies do not answer these questions, but lead to their
replacement. His work concentrates on visuo-spatial attention, which he
defines in terms of the following experimental paradigm. Posner studies
covert attentional effects in visual processing, that is, attentional effects
that are not facilitated by overt eye movements. A typical experiment involves
three locations on a screen facing an alert subject (Figure 9). In the
middle of the screen is a fixation 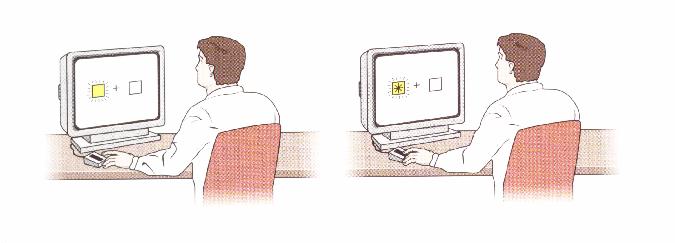
Figure 9. From Posner and Raichle (1994), p. 156.
point on which the subject must focus her vision. There is a square on each side of the fixation point, separated by some angle of eccentricity. In a typical experiment, the subject first fixates on the fixation point. Next, the subject is cued by a luminance increment in one of the boxes. The subject is trained not to respond to this luminance increment with a saccade or any other motoric response. Finally, a target, like an asterisk, appears in one of the boxes. The subject's task is to respond to the target, with a key press for example, as quickly as possible. Posner has obtained the following results with this experimental paradigm. If the target appears in the cued location within 100 msec after the cue, then the subject's processing of that information is facilitated in comparison with the condition where the target appears at an uncued location. This facilitation is measured in terms of reaction time. Thus, the subject takes longer to respond when the target appears at an uncued location than when it appears at the cued location.
Prima facie such results can be accounted for using the classical model of limited attentional resources. Visual space is represented by an automatic, pre-attentive, parallel cognitive resource. The representations of the different locations in this space compete for further processing by the limited attentional resource. The cuing of a particular location in visual space leads to the selection of the representation of that location for further processing. Assuming that a response to the target requires processing by the limited attentional resource, it is not surprising that responses to targets appearing in cued locations are facilitated. The cue has already led to the selection of that location for further, attentional processing, so when the target appears there it can be processed more rapidly than when it appears at a location that has not been cued, and is not already being processed by the limited attentional resource.
Despite its prima facie compatibility with the classical cognitive analysis
of attention in terms of selection for processing by a limited resource,
Posner develops a different and more complicated model of visuo-spatial
attention based on more fine-grained evidence obtained from experiments
with subjects with brain lesions and from PET studies. This work constitutes
an early example of neural evidence modifying and constraining the development
of a cognitive model. The PET studies show that a massively distributed
network of brain areas involving parietal cortex, the midbrain, and the
thalamus is active during performance of the covert, visuo-spatial attention
task (Posner, et al. 1994, p.221). And the lesion studies show that different
deficits in the performance of this task result from lesions in different
areas of this network.(ibid.) Posner uses such evidence to argue for the
claim that visuo-spatial attention is realized by elementary cognitive
operations, each of which is localized in a specific neural structure (Posner
1992, p.12). Because parietal lesions only affect the capacity to shift
attention from the ipsi- to the contralesional side, and not vice versa,
Posner argues that the posterior parietal lobe must be responsible for
disengaging attention to a location in the ipsilesional visual hemifield.
Because damage to midbrain structures (especially the superior colliculus)
affects the capacity to shift attention in both directions, Posner argued
that these midbrain structures must be responsible for the actual move
from one attended location to another. Analogous lesion evidence and similar
argumentation leads Posner to conclude that parts of the thalamus (the
pulvinar) are responsible for an engage operation, whereby attention to
a new location is engaged after a shift from an old location (ibid.). (Figure
10) 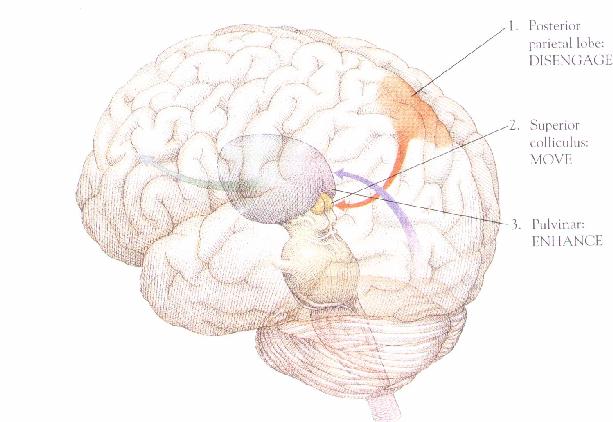 Fig. 10. From Posner and Raichle (1994), p. 168
Fig. 10. From Posner and Raichle (1994), p. 168
It is clear that Posner's fine-grained analysis of the role of specific neural structures in the performance of the covert, visuo-spatial attention task constitutes a radical reconceptualization of the cognitive role of attention. The metaphor of the limited processing bottle-neck is replaced by a "spotlight" metaphor (Posner, et al. 1994, p.224). These metaphors differ along several interesting dimensions. Posner's work suggests that attentional deficits are not deficits in types of processing, but rather deficits in efficiency of processing. Victims of lesions with attentional deficits still have the capacity to process information from every sensory modality, as well as semantic information, and high-level information about visual form (Posner, et al. 1984, p.1873). Their problem is that they cannot enhance this processing to the requisite degree when it is required for the purposes of some task. The spotlight metaphor is a more appropriate way of thinking about these deficits. It is not that some information cannot make it into a limited capacity bottle-neck. Rather, the system responsible for enhancing processing in primary sensory areas, as demanded by some task, is disabled with respect to one of these areas. The spotlight has trouble 'reaching' some areas of visual space. Along with this reconceptualization comes a cognitive/functional distinction that makes little sense on the traditional processing bottle-neck model of attention. When we reconceptualize attention as a spotlight, it makes sense to distinguish between the "site" and the "source" of attention (Corbetta, et al. 1991, p.2384) The site of attention is the neural area whose activity is enhanced for the purposes of some task. The source of attention is the neural area that effects this enhancement. Thus, where the traditional bottle-neck model of attention makes a distinction between two types of processing, i.e., pre-attentive vs. attentive, that take place at different sites, the spotlight model of attention sees attention as the enhancement of processing at various pre-attentive sites, caused by sources of enhancement like the posterior attention network.
The site/source distinction emerges quite naturally in the neurally informed study of attention. When one studies brain anatomy, no obvious candidates suggest themselves as locations of the processing bottle-neck that defines attention in the classical model. All one sees is massively parallel, distributed processing. For the neurally-informed student of attention, attention is not a separate, limited-capacity processing module. Rather, it is the activity of coordinating parallel processing streams through selective enhancement. As Allport argues, the only limited resource that parallel, pre-attentive processing streams compete for is the control of behavior (Allport 1989, pp.648-652). Coherent, coordinated behavior inevitably involves selectivity. Only a small number of overt actions can be performed at the same time. For example, one cannot both raise one's arm and lower it at the same time. The temptation to treat attention as a limited-capacity processing module is an artifact of the fact that the tasks used to measure attention involve coordinated, overt responses, and thus the inevitable selection of some processing streams over others. Thus, to speak of competition for a limited processing resource is less an explanation of attention than a description of what needs to be explained: the fact that in the type of coordinated activity that is required in tasks used to measure attention, only one processing stream can control overt behavior at a time (ibid., p.640). The 'spotlight' metaphor and the site/source distinction, on the other hand, suggest a functional analysis of attention that is much more explanatory, relative to facts about the brain.
Despite the reconceptualization of attention as a spotlight, many of the questions suggested by the bottle-neck model remain. It is still not obvious how selection works. On what basis does the spotlight select a specific neural area to enhance? Posner's work suggests that low level cues, like luminance changes at specific locations in the visual field, may lead to the enhancement of relevant neural areas. This seems to be an example of "stimulus-driven attentional capture" (see Yantis 1993). In both Posner's covert attention shift task, and the 'obvious' condition of the visual search task, attention is the product of a pre-attentive process. However, performance on the 'non-obvious' condition of the visual search task suggests a very different attentional effect. Here, the actual search seems to demand attention. Attention is part of the controlled, sequential processing that is necessary to locate the target. Posner tries to localize the division between the kind of attention that is automatically captured by luminance changes at locations in the visual field, and the kind of controlled attentional processing required for serial search, in an anatomical division between what he calls the anterior and posterior attention networks. As we have seen, the posterior attention network explains the capture of attention by luminance changes at specific locations in the visual field. Posner argues that the anterior attention network, and especially a neural area called the anterior cingulate, is responsible for the kind of controlled, attention-demanding processing that we see in tasks involving search for non-obvious targets. He hypothesizes that the anterior attention network (Figure 11) is where non-modality-specific, linguistically conveyed information is brought to bear on the control of attention (Posner, et al. 1994, p.219). Such information permits the controlled, attention-demanding search for targets which are not distinguished from distractors by obvious, modality-specific cues.
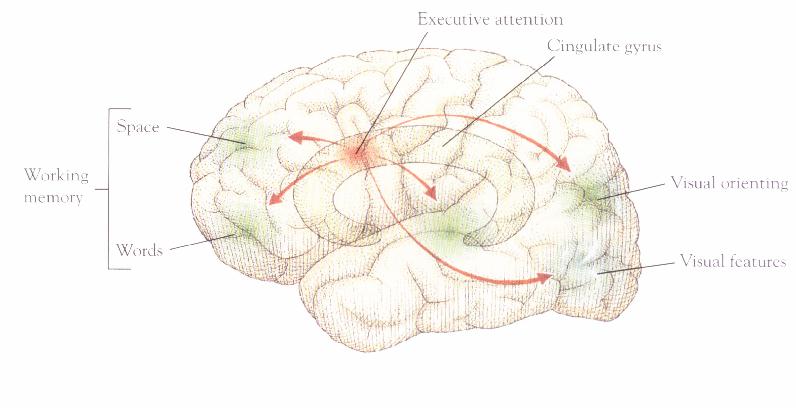
Figure 11. From Posner and Raichle (1994). p. 173.
However matters are not quite so straightforward. In a PET study, Corbetta, et al., examined the neural activation that accompanies performance on a visual search task involving two frames representing an array of moving objects. The subjects were asked to report if the two frames were the same or different with respect to certain visual features, like color, shape and speed. There were two conditions. In the 'divided-attention' condition subjects were asked to report if they noticed a difference in any of the object features, color, shape, or speed. In the 'selective-attention' condition, subjects were asked to report if they noticed a difference in a specific object feature, either color, shape, or speed. These two conditions yielded dramatic differences in the pattern of neural activation. Where the selective-attention condition involved primarily the enhancement of early visual areas responsible for the detection of the relevant feature, the more attention-demanding divided-attention condition involved additional activation of the anterior cingulate (Corbetta, et. al 1991). This seems to confirm that the anterior cingulate is involved in controlled, attention-demanding processing. However the experiment also seems to suggest that Posner's characterization of the difference between anterior attentional function and posterior attentional function is misleading, for in both the divided- attention and the selective-attention conditions, non-modality-specific, linguistic instructions play a role in the control of attention. In the selective-attention condition, which does not result in activation of the anterior cingulate, the instruction to detect changes in color as opposed to shape or motion, for example, activates the appropriate automatic selection for attention.
Corbetta et. al.'s results are consistent with increasing evidence from behavioral studies of attention that there are very few examples of stimulus information that can capture attention 'from the bottom up'. In other words, it is hard to specify what captures attention in a task- neutral way. Many high level features, like word form or even semantic properties, can capture attention automatically (Duncan 1980, p.291). Conversely, attention to supposedly compelling low level cues, like luminance changes, can be successfully and automatically inhibited if the task requires attention to properties that vary independently of such low level cues (Yantis 1993). Such evidence leads Steven Yantis to argue that Posner's studies are not examples of bottom-up attentional capture by luminance changes, since the requirements of the task, i.e., reaction to target onset, necessitate the detection of luminance changes (ibid., p.159). Given this argument, Posner's division of attention into a posterior system that is driven by visual stimuli and an anterior system that is driven by top-down, task-specific information seems ill-motivated.
If attentional capture by luminance increments at locations in the visual field is just a special case of automatic attentional capture by stimulus information that is most relevant to successful performance on the specific task that the subject is engaged in, then it may be better to think of such automatic attentional capture as a general phenomenon that can affect many different processing dimensions, rather than as a specific attentional system that scans visual space for inherently salient cues, like a spotlight. Such a reconceptualization still retains many features of the spotlight model. It still makes sense to make a distinction between the site and the source of attentional enhancement of processing. The difference is that there are many possible sites of such attentional enhancement besides the processing centers that represent visual space, and there may be correspondingly many sources of such attentional enhancement. Something like this model is suggested by Desimone and Duncan (1995, pp.199-200). They argue that attentional selection involves biased competition among different processing streams for the control of behavioral response. The competitive bias comes from an "attentional template" that is formed in conformity with task demands. In this model, the attentional template is the source of attentional enhancement, and visual areas that meet the requirements of the templates are the sites of attentional enhancement.
Desimone and Duncan's model has the advantage of offering both an explanation of automatic attentional capture by task-relevant features, and an explanation of why certain tasks require controlled, attention-demanding processing: when targets and distractors are not distinct along some obvious perceptual dimension, both representations of targets and of distractors get an equivalent competitive bias (ibid.). However, this model raises many new questions. Where exactly are attentional templates located? Are the attentional templates appropriate for different tasks located in distinct brain areas, or in the same brain area? How exactly is the weak competitive bias propagated from attentional templates to sensory processing areas? How are instructions translated into attentional templates? When targets and distractors are insufficiently distinct to yield automatic attentional capture, how and where does controlled, attention- demanding processing take place, and what is the role of templates in such processing? If attentional templates play no role in such controlled processing, then how do instructions yield appropriate sequences of controlled, attention-demanding processing?
Thus, there have been at least three cognitive/functional conceptualizations of attention, and both neural and cognitive psychological evidence have contributed to their articulation. Based primarily on behavioral evidence, attention was first conceptualized as a processing bottle-neck located somewhere between perception and motor response. Neural evidence threw doubt on this conceptualization. The brain's massively parallel processing makes the existence of such a bottle-neck unlikely. This led some to reconceptualize the limited resource that stimulus information seems to compete for as the control of overt behavioral response, and the selectivity that adjudicates such competition as the result of enhancement and amplification of specific processing sites. The sources of such enhancement and amplification were thought to be multiple. Some involved top-down task-specific information, while others seemed to involve low-level visuo-spatial cues. Posner identified these sources of attentional enhancement as the anterior and posterior attentional networks, respectively. However, behavioral evidence suggests that Posner's division is ill-motivated. Most, if not all, automatic, attentional capture, by supposedly obvious visual and sensory cues, is determined by top-down influences, i.e., by task-specific information. Thus, the distinction between a visual-cue based posterior attentional network, and a top-down task-specific anterior attentional network seems spurious. However, the distinction between automatic attentional capture, and target detection that requires controlled, attentional processing still seems robust. It is reasonable to suggest that the anterior attentional network, and especially the anterior cingulate, is crucial to the latter, while the former is performed by a diversity of attentional systems associated with relevant sensory areas, in something like the way proposed by Desimone and Duncan's biased competition, attentional template theory.
4. The Interactive Approach and the Dynamical Systems Challenge
Our goal has been to illustrate the interactive processes of functional decomposition and structural localization which is guiding the development of mechanistic models of cognitive phenomena in cognitive neuroscience. In this last section, though, we return briefly to the opposition to such research. We noted at the outset that there is a long tradition of opposition to the decomposition and localization of cognitive function. Flourens provided a strong opponent to Gall. Motivated largely by Cartesian considerations, but also influenced by lesion experiments which he performed on pigeon brains, Flourens argued that there was no specialization of function in the cerebral cortex:
All sensations, all perceptions, and all volition occupy concurrently the same seat in these organs. The faculty of sensation, perception, and volition is then essentially one faculty (1824, p. 241)
Lashley's opposition to cerebral localization was also inspired by lesion work, this time on rats, which showed no deficits to maze running ability after removal of large portions of the frontal cortex. Lashley (1929). Lashley interpreted this as evidence both for equipotentiality of frontal cortex tissue (the ability of any part to carry out functions of the whole part) and mass action (the degree of performance of a task depends upon how much of the original part remains). Lashley made exceptions to these principles for dedicated sensory and motor areas, but applied them to large parts of the cortex. He also suggested an interpretation of these principles in terms of Gestalt psychology's principle that higher level activities depend upon the dynamic organization of the entire cortex.
Lashley's appeal to Gestalt psychology's interest in dynamic organization links it to today's major opposition to decomposition and localization, stemming from some advocates of dynamical systems theory. Van Gelder (1995), for example, associates what he calls homuncularity with other concepts such as representation, computation, and sequential and cyclic operation, and contrasts this combined package with the dynamicist's view that seeks dynamical laws (taking the form, e.g., of differential equations) connecting different parameters in the overall system. It is reasonably clear that homuncularity is linked with notions of representation and computation.The notions of sequential and cyclic operations do not seem so intimately linked, since mechanists will often abandon sequential operations in their models when these provide inadequate accounts of the phenomena, and the notion of cyclic process van Gelder identifies, in which a system runs a complete cycle before receiving new input, is largely a product of modern computer design. In decomposing the cognitive system into different component functions, one identifies different information being supplied to each component, (hence different representations), and associates with each component a computation, where computation means some operation on the represented information. It is true that some dynamical models have a holistic, not decompositional character to them--variables are identified in the behavior of the system, and laws are sought relating these variables. But this task of discovering regularities in behavior is not incompatible with trying to explain the behavior of a system in terms of its components (Bechtel, 1996). The two tasks are complementary. Moreover, a decomposition and localization of cognitive function may provide the basis for identifying variables that figure in an dynamical analysis.
There is a different strand in the dynamicist's criticism, however, that does directly threaten the sort of decomposition and localization we have identified in cognitive neuroscience. This strand argues that the evidence usually adduced for decomposition and localization fails to demonstrate that there really are different parts doing different functions. van Orden and Stone (in press) argue that one of the strongest form of evidence presented for dissociation of function, double dissociation studies in which two patients show contrasting deficits and preservations of function, fail to do so. They generalize this criticism to interference studies of the sort we mentioned earlier and van Orden and Paap (in press b) advance a similar argument against PET studies. The basis for these objections is that the evidence that purports to show different functional components in different neural systems is subject to alternative interpretations in which a much more integrated system is damaged in different ways. Cohen et al. (1994), for example, develop a neuronal network model which simulates the normal and lesioned behavior of the disengage mechanism in Posner's account of the posterior attention system without including components whose specific task is to disengage attention. Other neural network studies such as Hinton and Shallice (1991) likewise show that lesions to interactive networks can show patterns of deficit resembling those one gets in human lesions without there being a component specifically responsible for the lost ability. Moreover, one can get a pattern of activation in a given area of a system without that component performing a distinctive sub-task of the overall activity.
This latter challenge from DST is a serious one. One way of viewing it, though, is as an instance of the more common phenomenon of the underdetermination of scientific theories. Any amount of empirical evidence is subject to alternative interpretations. The challenge in science is to develop the most plausible interpretation of evidence at a time and pursue research programs that build from that evidence to a more detailed and adequate account of the phenomenon. The question then becomes not whether the evidence adduced proves decomposition and localization, but whether it is part of a progressive research program. Cognitive neuroscience certainly seems to show all the signs of a being a progressive research program, and hence this is not the time to abandon it.
Footnotes
[1] A controversial issue, but one which does not affect the analysis we are offering here, is how one characterizes this input information. Many have adopted a static and atomized view of the input. This was severely criticized by Gibson (1974), who argued that the information that was picked up by a nervous system was far more integrated and dynamic. Gibson and many inspired by him have pointed out how information about time to impact with another object is provided through looming, for example--the speed of the spread of the pattern on the retina.
[2] Yet a third stream through LGN has been distinguished, a koniocelular (K) stream, but its limited cortical projection and sluggish response patterns led researchers to discount its significance in transmission of ascending visual information.
[3] Commenting on the methods of mapping, van Essen and DeYoe state: "The 32 distinct visual areas shown in this scheme have been identified on the basis of one or more of the following criteria: (1) a distinctive pattern of inputs and outputs; (2) a distinctive architecture, (3) a topographically organized representation of part or all of the contralateral visual hemifield; and (4) a characteristic functional signature revealed by physiological recordings or restricted cortical lesions" (p. 386).
[4] When the connections are between lateral areas, the reciprocal connections tend to be symmetrical--connections in both directions start in the same layer and terminate in the same layer. This is not true when the connections are within a processing stream.
References
Allport, D.A. (1989). "Visual Attention," in M. I. Posner (Eds.), Foundations of Cognitive Science (pp.631-682). Cambridge MA: MIT Press.
Bechtel, W. (1986). Building interlevel pathways: The discovery of the Embden-Meyerhof pathway and the phosphate cycle. In J. Dorn and P. Weingartner (Eds.) Foundations of biology, pp. 65-97. Vienna: Holder-Pichler-Tempsky.
Bechtel, W. (1996). Yet another revolution? Defusing the dynamical system theorists' attack on mental representation. Presidential Address to the Society for Philosophy and Psychology.
Bechtel, W. and Richardson, R. C. (1993). Discovering complexity: Decomposition and localization in scientific research. Princeton: Princeton University Press.
Broadbent, D. E. (1958). Perception and Communication. London: Pergamon.
Broca, P. P. (1861). Remarques sur le siège de la faculté du langage articulé; suivies d'une observation d'aphémie (perte de la parole). Bulletin de la Société de l'Anatomie de Paris, 36, 330-357.
Chalmers, D. (1996). The conscious mind. Oxford: Oxford University Press.
Chomsky, N. (1980). Rules and representations. The Behavioral and Brain Sciences, 3, 1-15.
Clark, E. and Jacyna, L. S. (1987). Nineteenth century origins of neuroscientific concepts. Berkeley: University of California Press.
Cohen, J. D., Romero, R. D., Servan-Schreiber, D. and Farah, M. J. (1994). Mechanisms of spatial attention: The relation of macrostructure to microstructure in parietal neglect. journal of Cognitive Neuroscience, 6, 377-387.
Corbetta, M. Miezin, F. M., Dobmeyer, S., Shulman, G. L. and Petersen, S. E. (1991). Selective and divided attention during visual discriminations of shape, color, and speed: Functional anatomy by positron emission tomography, Journal of Neuroscience, 11 (8): 2383-2402.
Craig, B. G. (1969). The topography of the afferent projection in the circumstriate visual cortex (C.V.C.) of the monkey studied by the Nauta method. Vision Research, 9, 733.
Dennett, D. C. (1978). Brainstorms. Cambridge, MA: MIT Press
Desimone, R. and Duncan, J. (1995). Neural Mechanisms of Selective Visual Attention,.Annual Review of Neuroscience, 18: 193-222.
Donders, F. C. (1868/1969). On the speed of mental processes. Acta Psychologica, 30, 412-431.
Duncan, J. (1980). The locus of interference in the perception of simultaneous stimuli. Psychological Review, 87 (3): 272-300.
Felleman, D. J. and van Essen, D. C. (1991). Distributed hierarchical processing in the primate cerebral cortex. Cerebral Cortex, 1, 1-47.
Flourens, J. P. M. (1824). Recherches Expérimentales sur les Propriétés et les Fonctions du Système Nerveux dans les Animaux Vertébrés. Paris: Crevot
Fodor, J. A. (1974). Special sciences (Or: Disunity of science as a working hypothesis. Synthese, 28, 97-115.
Fodor, J. (1980). The modularity of mind. Cambridge, MA: MIT Press.
Gall, F. J. (1825). Revue critique de quelques ouvrages anatomico-physiologiques, et exposition d'une nouvelle philosophie des qualités morales et des facultés intellectuelles. Volume 6 of Sur les fonctions du cerveau et celles de chacune de ses parties, avec des observations sur la possibilité de reconnaitre les instincts, les pechans, les tallens, ou les dispositions morales et intellectuelles des hommes et des animaus, par la configuration de leur cerveau et du leur tête. Paris: J. B. Baillière. English translation in 1835 by Winslow Lewis, Jr. Boston: Marsh, Capen, & Lyon.
Gall, F. J. (1812). Anatomie et physiologie du systême nerveaux et général, et du cerveau en particulier, avec des observations sur la possibilité de reconnoitre plusieurs dispositions intellectuelles et morales de l'homme et des animaux, par la configuration de leur têtes. Volume 2. Paris: F. Schoell.
Geschwind, N. (1974). Selected papers on language and brain. Dordrecht: Reidel.
Gibson, J. J. (1966). The senses considered as perceptual systems. Boston: Houghton Mifflin.
Hale, S., Chen, J., Myerson, J. and Simon, A. (1996). Behavioral evidence for brain-based ability factors in visuospatial information processing. Paper presented at the Psychonomics Society, November.
Hale, S. and Jansen, J. (1994). Global processing-time coefficients characterize individual and group differences in cognitive speed. Psychological Science, 5, 384-389.
Haxby, J. V., Grady, C. L., Horwitz, B., Ungerleider, L. G., Mishkin, M., Carson, R. E., Herscovitch, P., Schapiro, M. B., and Rapoport, S. I. (1991). Dissociation of object and spatial visual processing pathways in human extrastriate cortex. National Academy of Sciences, USA, 88, 1621-1625.
Hinton, G. and Shallice, T. (1991). Lesioning a connectionist network: Investigations of acquired dyslexia. Psychological Review, 98, 74-95
Jacobs, R. A., Jordan, M. I., and Barto, A. G. (1991). Task decomposition through competition in a modular connectionist architecture: The what and where visions tasks. Cognitive Science, 15, 219-250.
Lashley, K. S. (1929). Brain mechanisms and intelligence. Chicago: University of Chicago Press.
Lycan, W. G. (1981). Form, function, feel. Journal of Philosophy, 78, 24-49.
Macko, K. A., Kennedy, C. Sokoloff, L. and Mishkin, M. (1981). Society for Neuroscience Abstracts, 7, 832.
Marr, D. (1982). Vision: A computational investigation into the human representation and processing of visual information. San Francisco: W. H. Freeman.
Maunsell, J. H. R. and Newsome, W. T. (1987). Visual processing in monkey extrastriate cortex. Annual Review of Neuroscience, 10, 363-402.
McCauley, R. N. (1996). Explanatory pluralism and the co-evolution of theories in science. In R. N. McCauley (Ed.) The Churchlands and their critics. Oxford: Basil Blackwell.
Mishkin, M., Ungerleider, L. G., and Macko, K. A. (1983). Object vision and spatial vision: Two cortical pathways. Trends in Neurosciences, 6, 414-417.
Mundale, J. and Bechtel, W. (1996). Integrating neuroscience, psychology, and evolutionary Biology through a teleological conception of function, Minds and Machines, 6.
Nealey, T. A., Ferrera, V. P., and Maunsell, J. H. R. (1991). Magnocellular and parvocellular contributions to the ventral extrastriate cortical processing stream. Society for Neuroscience Abstracts, 17, 525
Neisser, U. (1989). Direct perception and recognition as distinct perceptual systems. Address presented at the Eleventh Annual Meeting of the Cognitive Science Society, Ann Arbor, MI.
Pinker, S. (1994). The language instinct. New York: W. Morrow and Co.
Pohl, W. (1973). Journal of Comparative Physiology and Psychology, 82, 227-239.
Posner, M. I. (1992). Attention as a Cognitive and Neural System. Current Directions in Psychological Science, 1 (1): pp.11-14.
Posner, M. I.,. (1984). Effects of Parietal Injury on Covert Orienting of Attention. Journal of Neuroscience, 4(7): 1863-1874.
Posner, M.I., Grossenbacher, P. G., and Compton, P. E. (1994). Visual Attention. In M. Farah & G. Ratcliff (Eds.), The Neuropsychology of High-Level Vision: Collected Tutorial Essays, pp.217-239.
Putnam, H. (1975). Philosophy and our mental life. In H. Putnam (Ed.) Mind, language, and reality: Philosophical papers of Hilary Putnam (Volume 2, pp. 215-271). Cambridge, Cambridge University Press.
Rueckl, J. G., Cave, K. R., and Kosslyn, S. M. (1989). Why are "what" and "where" processed by separate cortical visual systems? A computational investigation. Journal of Cognitive Neural Science, 1, 171-186.
Spurzheim, J. C. (1818). Observations sur la phraenologie, ou la connaissance de l'homme moral et intellectuel, fondée sur les fontions du système nerveux. Paris: Treuttel & Wurtz.
Sternberg, S. (1967). Two operations in character recognition: Some evidence from reaction time experiments. Perception and Psychophysics, 2, 45-53.
van Essen, D. C. and DeYoe, E. A. (1995). Concurrent processing in the primate visual cortex. In M. S. Gazzaniga, ed., The Cognitive Neurosciences, pp. 383-400. Cambridge, MA: MIT Press.
van Gelder, T. (1995). What might cognition be, if not computation? The Journal of Philosophy, 92, 345-381.
van Orden, G. C. and Paap, K. R. (in press). Functional neuroimages fail to discover pieces of the mind in the parts of the brain. Philosophy of Science
van Orden, G. C., Pennington, B. F., Stone, G. O, (1996). What do double dissociations prove? Inductive methods and isolable systems. Manuscript.
Wernicke, C. (1874). Der Aphasiche Symptomcomplex: Eine Psychologische Studie auf Anatomischer Basis. Breslau: Cohen und Weigert.
Yantis, Steven (1993). Stimulus-Driven Attentional Capture. Current Directions in Psychological Science, 2 (5), 156-161.
Zeki, S. M. (1976). The functional organization of projections from striate to prestriate visual cortex in rhesus monkey. Cold Spring Harbor Symposium on Quantitative Biology, 40, 591-600.